研究概要
ヒトを含めた動物は日常で起こった出来事や学習によって得られた知識を生涯にわたり覚えておくことができます。このような記憶が体のどこにどのように保存されているか、ということは古代から多くの人々の関心の対象でした。現在では記憶情報が脳に保存されているということは広く知られていますが、脳の中のどの部分にどのような仕組みで保存されているのか等、詳細についてはまだ多くが不明のまま残されています。
私たちの研究室では、記憶が脳の 「どの部分」に、「いつ」、「どのような仕組みで」保存され、それがどのような機構で呼び出されるのかについて、細胞・分子レベルで解明することを目標として研究を行っています。具体的には、実験動物を研究対象として分子生物学、生化学、生理学、光学、動物行動学などのさまざまな分野の最新の手法を組み合わせて、記憶を形成・維持する際に大脳の神経細胞や神経回路の性質がどのように変化するのかを実験によって明らかにしていきます。
また、学習・記憶のみならず、情動や意思決定、認知的柔軟性なども含む認知機能全般の神経機構、さらには”意識”がどのように大脳で作られているのか、など個々の研究者の興味に沿った研究を展開しています。
現在進行中の研究テーマ
1. 活動依存的遺伝子 Arc の機能解析 [奥野]
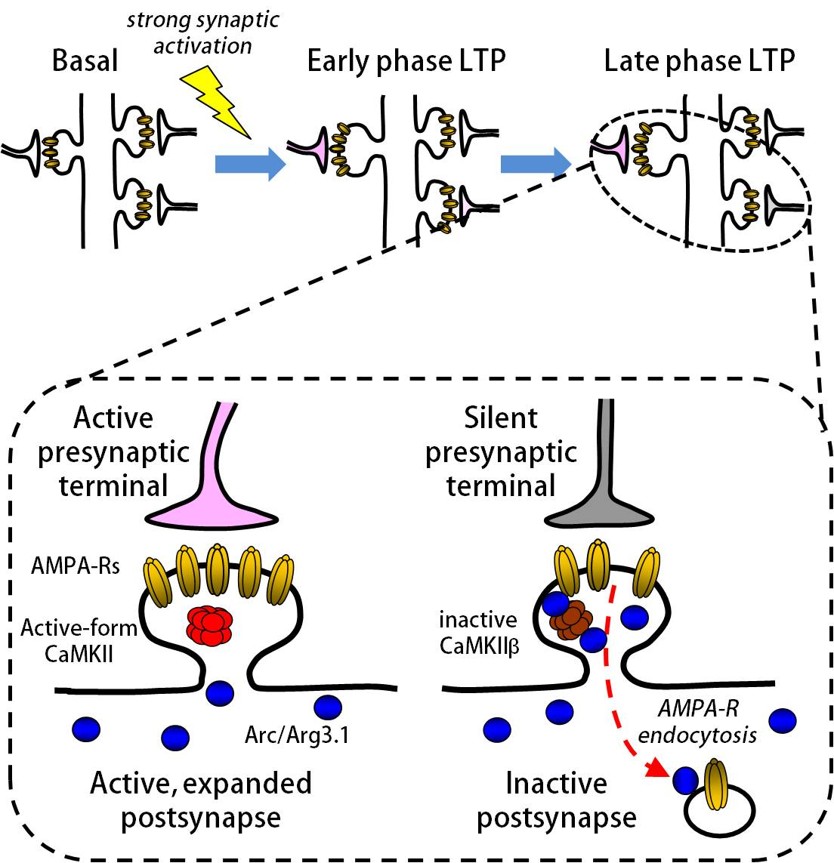
代表的な神経活動依存的遺伝子である Arc(Activity-regulated cytoskeletal-associated protein、別名 Arg3.1)はシナプス結合強度の調節に関与していることが知られています。私たちはこれまでに、Arc タンパク質が後シナプス部位でカルシウム・カルモジュリンキナーゼII(CaMKIIβ)と結合することにより局在が制御され、さらにグルタミン酸受容体のシナプス発現量を調節していることを明らかにしてきました(Chowdhury et al., Neuron, 2006; Okuno et al., Cell, 2012)。遺伝子改変マウスを用いた研究ではArc遺伝子が長期記憶に重要な役割を果たしていることが明らかになっていますが、その詳細な機構は不明です。現在、脳のどの部分でいつArcが発現することが記憶の形成や維持に必要なのかについて調べています。
2. 記憶関連大脳神経回路の可視化・同定 [奥野、城山]
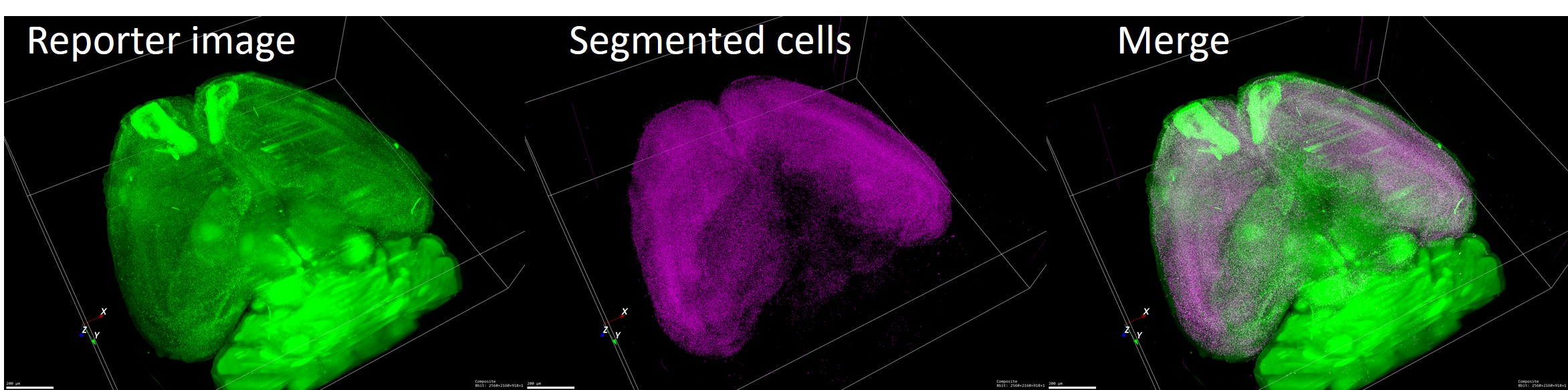
記憶を形成する際には大脳のすべての神経細胞が関与しているのではなく、特定の記憶に対してある特定の神経細胞群が記憶回路として動員されると考えられています。この神経細胞群を可視化することにより、大脳のどの部分に記憶が形成・維持されているのかを研究しています。私たちはこれまでに活動依存的なプロモーターを用いた蛍光レポーターシステムなどを開発してきました(Kawashima et al., PNAS, 2009; Kawashima et al., Nat Methods, 2013; Mikuni et al., Neuron, 2013; Vousden et al., Brain Struct. Funct., 2014)。現在、これらの技術を応用した神経活動レポーターマウスを作成し、組織透明化技術と組み合わせることにより全脳レベルでの網羅的な記憶神経回路の同定をめざしています。
3. 新規シナプス機能調節遺伝子の探索 [奥野、原口]
神経活動によって一過的に発現誘導される遺伝子は、上記の Arc の他にも c-fos、fosB、egr-1(zif268/NGFI-A)、Nr4a1(NGFI-B/Nur77)など様々なものが知られていますが、一部の遺伝子を除いて解析は進んでおらず、多くの遺伝子の機能は未知のままです。まだ解析が進んでいない活動依存的な遺伝子を調べることにより、新たなシナプス機能調節因子を探索しています。
4. 新たな認知課題の開発 [城山、鈴木]
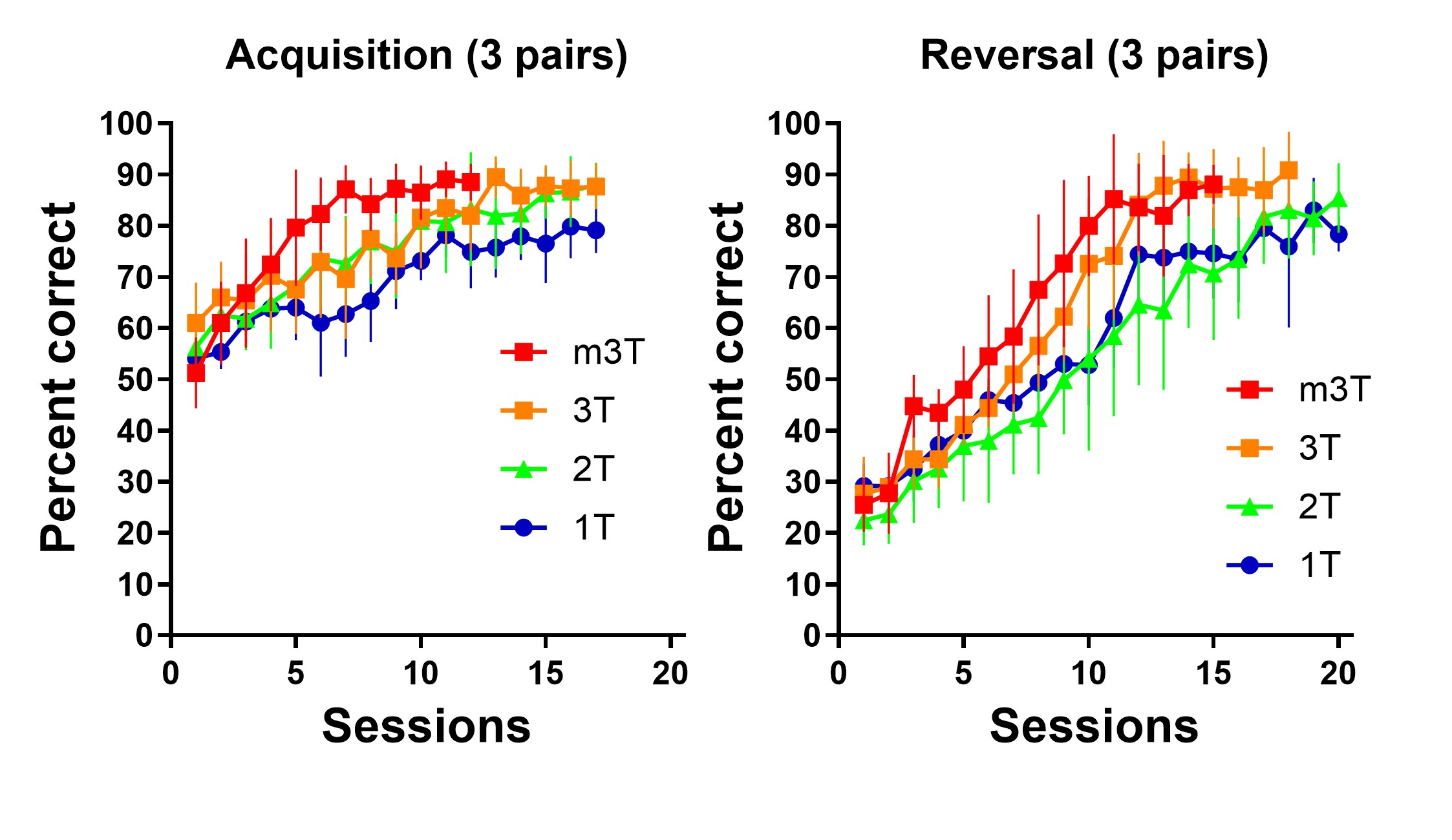
学習や記憶を含む認知機能をマウスなどの実験動物で調べるためには、その目的に適した行動課題が必要です。私たちの研究室では記憶や情動をテストするための古典的な行動課題以外にも、タッチパネルを用いたオペラント学習装置を用いて新規の行動課題や学習プロトコールを開発しています。
5. 大脳皮質-視床のネットワークの機能解明 [倉本]
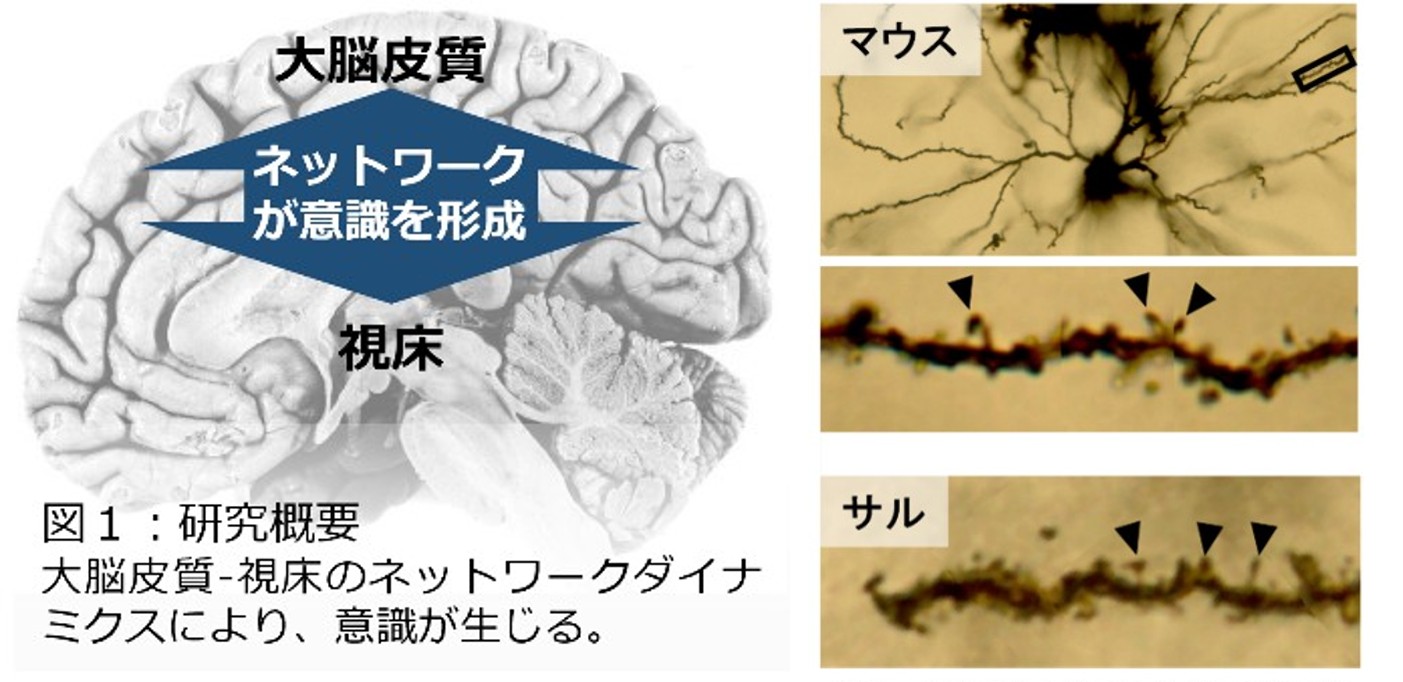
これまで視床は外界からの情報を大脳皮質に伝えるという中継地としての役割が主であると思われていました。しかしながら、視床が大規模に傷害されると、大脳皮質には問題がなくても、意識障害を生じます。このことから、”意識を生み出すためには大脳皮質-視床の相互作用が重要である”と考えられ、最近では視床の新しい機能が注目されています。私たちは視床の神経細胞には大脳皮質の神経細胞と同様に樹状突起に棘(スパイン)構造存在していることを見出し、この機能解析を進めています。
主要参考文献
- Chowdhury S, Shepherd JD, Okuno H, et al. Arc interacts with the endocytic machinery to regulate AMPA receptor trafficking. Neuron, 42:445–459 (2006).
- Kawashima T, Okuno H, et al. A synaptic activity-responsive element in the Arc/Arg3.1 promoter essential for synapse-to-nucleus signaling in activated neurons. Proc. Natl. Acad. Sci. USA, 106:316–321 (2009).
- Okuno H. Regulation and function of immediate-early genes in the brain: beyond neuronal activity markers. Neuroscience Res. 69:175–186 (2011).
- Okuno H, Akashi K, Ishii Y, et al. Inverse synaptic tagging of inactive synapses via dynamic interaction of Arc/Arg3.1 with CaMKIIβ. Cell, 149:886–898 (2012).
- Kawashima T, Kitamura K, Suzuki K, et al. Functional labeling of neurons and their projections using the synthetic activity-dependent promoter E-SARE. Nat Methods, 10:889–895 (2013).
- Kiyama Y, Suzuki Y, Haraguchi M, et al. A simple method that doubles learning speed for mice in touchscreen-based visual discrimination. Sci Rep, 15: 43133 (2025).